
En una entrada anterior introdujimos las nociones básicas acerca de los hidratos de carbono y su papel en la alimentación del ser humano. Continuando con este paseo por el mundo de los glúcidos pasamos ahora a hablar acerca de su metabolismo y del funcionamiento de algunas hormonas implicadas él. Este post contiene una dosis algo densa de bioquímica y fisiología que no agradará a algunos, pero creo que es importante conocer estos aspectos de cara a entender el papel de los hidratos de carbono en la nutrición
Reacciones catabólicas de hidratos de carbono
Denominamos reacciones catabólicas a aquellas que tienen por objeto convertir moléculas complejas en otras más sencillas, obteniendo energía en el proceso. Vamos a centrarnos en las principales reacciones o procesos que engloban a los hidratos de carbono.
Digestión de hidratos de carbono de los alimentos
La digestión no constituye en sí misma una reacción catabólica, pero es imprescindible para absorber los hidratos de carbono presentes en los alimentos y poder disponer de ellos, por lo que la vamos a comentar brevemente. Los hidratos de carbono pueden encontrarse en la alimentación en diversas formas, aunque para facilitar la comprensión de los procesos de digestión y absorción vamos a distinguir cuatro tipos:
- Monosacáridos: son las moléculas más simples, principalmente glucosa, fructosa y galactosa.
- Disacáridos: formados por dos moléculas de glúcidos, por ejemplo la sacarosa, la lactosa y la maltosa.
- Polisacáridos: formados por múltiples unidades de monosacáridos, a destacar amilosa y amilopectina (componentes del almidón vegetal).
- Fibra: conjunto de moléculas de distinta longitud que no pueden ser digeridas ni absorbidas, como la celulosa o el almidón resistente.
A pesar de la amplia variabilidad de estructuras, los monosacáridos glucosa, fructosa y galactosa son los únicos glúcidos con buena absorción intestinal, por lo que disponemos de distintos mecanismos para convertir cualquier polisacárido a estas estructuras más simples.
El primer proceso es la digestión del almidón, que comienza en la boca por medio de la α-amilasa salival. Esta enzima actúa sobre los enlaces α-1-4 de la amilosa y la amilopectina cortando las largas cadenas de almidón en otros componentes de menor longitud (maltosa, maltotriosa y α-dextrina límite), pero no tiene actividad sobre los enlaces α-1-6 (ramificaciones) ni sobre los β-1-4 de la celulosa (por eso ésta no puede ser digerida). La α-amilasa salival se inactiva por el pH ácido del estómago por lo deja de tener acción cuando aún quedan moléculas de almidón por digerir. El páncreas produce α-amilasa de acción idéntica a su homóloga salivar pero con mayor tasa de actividad.

Los oligosacáridos que quedan tras los procesos de digestión del almidón así como otros glúcidos que han sido ingeridos llegan hasta duodeno y yeyuno, donde han de ser absorbidos. Allí, las células del epitelio intestinal cuentan con enzimas oligosacaridasas, que completan los procesos de digestión de los glúcidos hasta convertirlos en monosacáridos para ser absorbidos. Existen tres oligosacaridasas principales: maltasa, dextrinasa (o isomaltasa) y lactasa; y los productos de su acción son tres monosacáridos: glucosa, fructosa y galactosa.
Quedan sin digerir los componentes de la fibra alimentaria, algunos de los cuales al llegar al colon son parcialmente metabolizados por las bacterias de la flora intestinal dando lugar a ácidos grasos de cadena corta, que son absorbidos y sirven de energía para las células del intestino.
Glucolisis
La glucolisis es la vía de metabolización de la glucosa hasta ácido pirúvico para obtener energía en forma de ATP (adenosina trifosfato) y NADH (dinucleótido reducido de nicotinamida y adenina) a través de una secuencia de diez reacciones enzimáticas. Es considerada la reacción (o más propiamente hablando serie de reacciones) central del metabolismo de los hidratos de carbono, pues en sus distintas etapas pueden acceder otros hidratos de carbono (galactosa, fructosa).
La glucolisis es uno de los procesos metabólicos más antiguos conocidos. Puede ocurrir en presencia o ausencia de oxígeno (aerobiosis y anaerobiosis respectivamente), y está presente en la práctica totalidad de organismos vivos, desde las archeobacterias hasta el ser humano, con mínimas variaciones. Se especula que incluso pudiera haberse producido de forma espontánea en el océano durante el Eón Arcaico, hace 4000 millones de años.

La glucolisis tiene varios puntos de regulación, lo que indica que distintas moléculas intermediarias pueden acceder al ciclo en diferentes etapas, y que es posible incrementar o reducir la velocidad a la que se lleva a cabo en función de las necesidades.
Así por ejemplo, la glucosa 6-fosfato actúa como indicador de los niveles de glucemia, ejerciendo una importante regulación; si estos son elevados la glucosa 6-fosfato bloquea la hexoquinasa (1) y en lugar de proseguir con la glucolisis hasta piruvato puede pasar a convertirse en glucosa 1-fosfato y pasar a otra vía para la síntesis de glucógeno. La glucosa es el único carbohidrato de origen alimentario que es sustrato de la hexoquinasa, por lo que la ingesta de fructosa u otros monosacáridos como endulzantes "saltan" este punto de regulación. Los niveles de energía, en forma de ATP, regulan la actividad de otras dos enzimas, la fosfofructo quinasa (3) y la piruvato quinasa (10). Estas tres reacciones son irreversibles.
El balance final de la glucolisis es el siguiente:
Glucosa + 2 NAD+ + 2 ADP + 2 Pi → 2 Piruvato + 2 NADH + 2 H+ + 2 ATP + 2 H2O
El ácido pirúvico puede, a partir de aquí, tomar diversas vías, entrando al ciclo de Krebs para continuar proporcionando energía, ser fermentado hasta ácido láctico, reconvertirse en glucosa vía gluconeogénesis, servir para la síntesis de ácidos grasos, convertirse en el aminoácido alanina o incluso en etanol.
El ácido pirúvico puede, a partir de aquí, tomar diversas vías, entrando al ciclo de Krebs para continuar proporcionando energía, ser fermentado hasta ácido láctico, reconvertirse en glucosa vía gluconeogénesis, servir para la síntesis de ácidos grasos, convertirse en el aminoácido alanina o incluso en etanol.
Fructolisis
La fructosa, a diferencia de la glucosa, es metabolizada casi completamente en el hígado, siendo su metabolismo en otros células reducido o inexistente. La fructosa es transformada en fructosa 1-fosfato por la enzima fructoquinasa, y se escinde por acción de la aldolasa en gliceraldehído y dihidroxiacetona fosfato. A través de distintas reacciones, estas moléculas pueden seguir las siguientes vías:
- Síntesis de glucógeno. Es la vía preferente de destino de la fructosa, si bien está limitada por la capacidad del hígado para rellenar sus reservas de glucógeno.
- Integración en la glucolisis y oxidación a piruvato. La dihidroxiacetona fosfato y el gliceraldehído pueden ser convertidos a gliceraldehído 3-fosfato, un intermediario de la glucolisis y la gluconeogénesis.
- Formación de acetil Coenzima A y síntesis de triglicéridos.
Glucogenolisis
El glucógeno es la molécula polisacarídica en la que las células almacenan unidades de glucosa. Es un polímero ramificado, similar a la amilopectina de los almidones vegetales y contiene los dos tipos de enlaces que ya hemos visto:
- α-1-4: enlaces lineales.
- α-1-6: enlaces de las ramificaciones; aparecen en mayor proporción que en las amilopectinas.

La enzima glucógeno fosforilasa rompe un enlace terminal de la molécula de glucógeno dando una molécula de glucosa 1-fosfato, la cual es convertida a glucosa 6-fosfato por la enzima fosfoglucomutasa y se integra en la glucolisis. Sobre los enlaces α-1-6 actúa otra enzima denominada glucosidasa, que produce glucosa.
Reacciones anabólicas de hidratos de carbono
Denominamos reacciones anabólicas a aquellas que tienen por objeto sintetizar moléculas complejas a partir de otras más sencillas, consumiendo energía en el proceso.
Gluconeogénesis
Ciertos tipos celulares, como los eritrocitos o algunas células del sistema nervioso carentes de mitocondrias solo pueden usar glucosa como molécula dadora de energía. Dado que las reservas de glucógeno del organismo son limitadas y no podemos depender continuamente de alimentos, contamos con vías metabólicas capaces de fabricar glucosa a partir de otras moléculas:

- Piruvato: su conversión en glucosa sigue un camino en sentido opuesto al de la glucolisis y hasta cierto punto paralelo, aunque varían las tres reacciones que, como comentamos anteriormente, son irreversibles en la glucolisis, y que en la gluconeogénesis emplean enzimas distintas.
- Lactato: el lactato puede convertirse en piruvato por acción de la lactato deshidrogenasa.
- Aminoácidos glucogénicos: todos los aminoácidos con la excepción de la leucina y la lisina pueden dar lugar a glucosa previa conversión a piruvato o a alguna de las moléculas intermediarias del ciclo de Krebs.
- Glicerol: el glicerol es una molécula de tres átomos de carbono que forma parte de los triacilgliceroles, fosfolípidos y otras moléculas lipídicas, y que puede ser convertido en gliceraldehído 3-fosfato, un intermediario de la glucolisis y la gluconeogénesis.
2 Piruvato + 2 NADH + 2 H+ + 4 ATP + 2 GTP + 2 H2O → Glucosa + 2 NAD+ + 4 ADP + 2 ADP + 6 Pi
Si se compara la energía producida en la glucolisis con la consumida en la gluconeogénesis salta a la vista que es un proceso energéticamente muy costoso, y que se gasta más energía en recomponer una molécula de glucosa que en romperla. Por eso, se entiende que esta vía debe ser imprescindible para garantizar la viabilidad de ciertas células, y que no se va a llevar a cabo salvo que el balance total de procesos del metabolismo resulte favorable.
Glucogenogénesis
Se denomina glucogenogénesis al proceso de síntesis de una molécula de glucosa a partir de un precursor monosacarídico, la glucosa 6-fosfato. Las moléculas de glucosa 6-fosfato son convertidas en glucosa 1-fosfato, que forma cadenas con enlaces α-1-4. Posteriormente se pueden formar ramificaciones con enlaces α-1-6 por adición de cadenas más pequeñas a la cadena principal.
La gluconeogénesis tiene lugar principalmente en el hígado y el tejido muscular, y está sometida a un estrecho control hormonal.
Insulina, hormonas maestra del metabolismo de los hidratos de carbono
La gluconeogénesis tiene lugar principalmente en el hígado y el tejido muscular, y está sometida a un estrecho control hormonal.
Insulina, hormonas maestra del metabolismo de los hidratos de carbono
Todos los procesos anteriormente comentados, así como otros que hemos dejado de nombrar por su menor interés, están finamente regulados por distintas hormonas, las cuales dirigirán el balance global del metabolismo de los hidratos hacia el anabolismo o el catabolismo en función de las necesidades del organismo. Hablar del metabolismo de los hidratos de carbono nos lleva ineludiblemente a hablar de la insulina.
Efectos de la insulina
La insulina es una hormona polipeptídica producida por las células β de los islotes pancreáticos. Tras la ingesta de hidratos de carbono en una comida se produce una elevación de las concentraciones de insulina en sangre, que provoca una rápida captación de estos hidratos de carbono por los tejidos, induciendo su almacenamiento y utilización apropiados.
La insulina es una hormona polipeptídica producida por las células β de los islotes pancreáticos. Tras la ingesta de hidratos de carbono en una comida se produce una elevación de las concentraciones de insulina en sangre, que provoca una rápida captación de estos hidratos de carbono por los tejidos, induciendo su almacenamiento y utilización apropiados.

La insulina facilita la captación de glucosa en el músculo y el hígado, favorece la acción de las enzimas de la glucogenogénesis e inhibe la gluconeogénesis. Además, si la cantidad de carbohidratos ingeridos supera la capacidad de las células hepáticas la insulina induce su conversión a ácidos grasos, que serán posteriormente transportados hasta el tejido adiposo; aumenta la captación de glucosa por los adipocitos e impide la hidrólisis de los triglicéridos por inhibición de la lipasa sensible a insulina. También tiene influencia sobre el metabolismo de las proteínas, favoreciendo la captación de aminoácidos por las células, aumentando la síntesis de proteínas e inhibiendo su degradación.
En términos globales podemos decir que la insulina fomenta el empleo de hidratos de carbono como fuente de energía a la vez que impide el uso de los lípidos. Si la glucemia se eleva se secreta insulina, los hidratos de carbono se utilizan con fines energéticos y su exceso se almacena en forma de glucógeno y grasa; si la glucemia desciende, las células utilizarán las grasas como fuente de energía. Pero además, es una hormona imprescindible para el correcto metabolismo de grasas y proteínas.
Mecanismo de acción de la insulina y alteraciones
Ante una comida con una gran cantidad de hidratos de carbono se libera una gran cantidad de insulina con el objetivo de regular la glucemia sanguínea. La rápida llegada de insulina al torrente sanguíneo permite la entrada de glucosa en las células para ser utilizada como combustible y almacenarse en forma de glucógeno. Llegados a este punto es necesario aclarar (que no desmentir) un mito bastante extendido: "el exceso de carbohidratos de almacena en forma de grasa". Es cierto, como hemos explicado, que una vez se ha superado la capacidad del hígado para almacenar glucógeno, la glucosa puede transformarse en ácidos grasos (lipogénesis) para su reserva en el tejido adiposo; sin embargo, este es un proceso relativamente lento, necesitándose ingestas muy abundantes e incluso más de una comida para que comience este proceso. Facilitan la lipogénesis algunos factores, como la dieta alta en hidratos de carbono de forma crónica (pues los depósitos de glucógeno ya estarán llenos previamente) o el consumo de altas cantidades de fructosa (que solo puede almacenarse como glucógeno hepático pero no muscular).
Ya hemos visto que la insulina se secreta con el objetivo principal de controlar los niveles de glucemia, pero ni son los carbohidratos el único macronutriente que estimula la secreción de insulina, ni la función de ésta es únicamente el manejo de los hidratos de carbono. Comidas que contienen baja cantidad de hidratos de carbono también provocan la secreción de insulina, concepto importante y que tenemos que tener claro para no caer en la falsa creencia de que "la insulina nos hace engordar". Se han estudiado los efectos de un gran número de alimentos sobre la secreción de insulina, descubriéndose que algunas sustancias como la carne de res, el suero de leche (whey), o incluso el aminoácido leucina provocan elevaciones en la secreción de insulina que pueden llegar a alcanzar la misma magnitud que se produciría con el consumo de hidratos de carbono.

Parece entonces poco lógico decir que la insulina es causante de obesidad o que que mantener niveles bajos de insulina nos permitiría adelgazar, y que el consumo crónico y por encima de los límites de tolerancia de cada persona de hidratos de carbono juega un papel más importante en este fenómeno.
En individuos sanos existen dos ritmos de secreción de insulina que se superponen: uno basal, que regula constantemente los niveles de glucemia; y uno en forma de picos como respuesta a las comidas, que facilita la utilización de los nutrientes ingerido; la producción de estos picos de insulina es una situación fisiológica, y es imprescindible para la correcta regulación del metabolismo energético. En personas sanas se repite un ciclo constante de hiperglucemia - hiperinsulinemia - hipoglucemia reactiva a lo largo del día, cuyas oscilaciones serán tanto más pronunciadas conforme mayor sea la cantidad de hidratos de carbono que se consume.
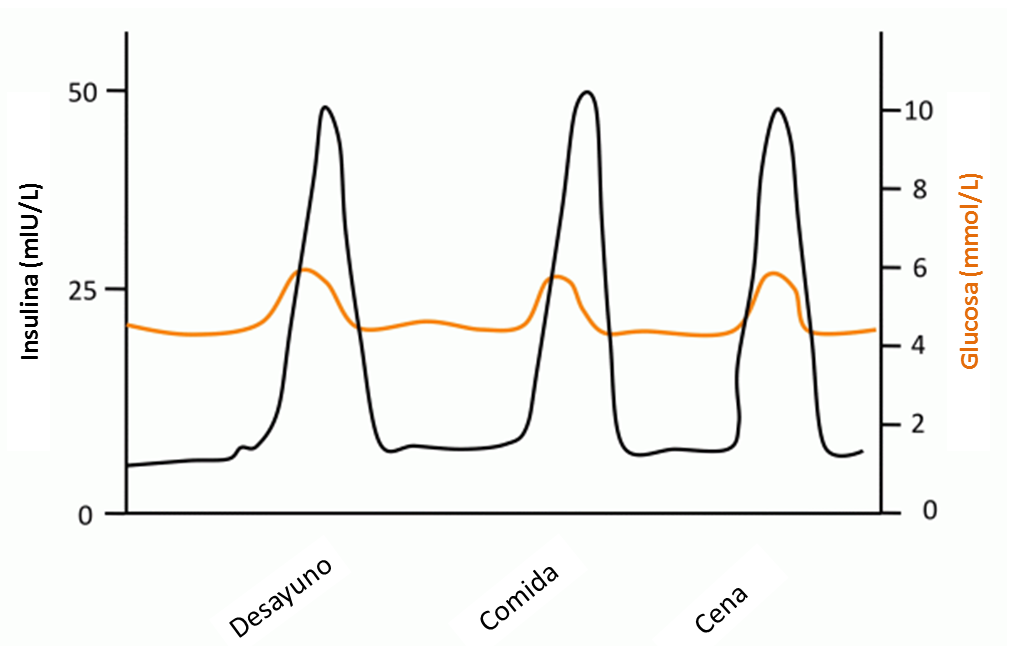 Cuando se produce una ingesta excesiva a lo largo de los años tiene lugar una serie de consecuencias negativas; así, por ejemplo, las grandes hiperglucemias seguidas de hipoglucemias posprandiales pronunciadas alteran los ritmos de hambre y saciedad, obligando a comer continuamente y desarrollando un apetito dirigido a los productos de mayor densidad calórica y altos en hidratos de carbono refinados. La conjunción de éste y otros factores (sedentarismo, obesidad) dan lugar a alteraciones a nivel celular (reducción de transportadores de glucosa GLUT4, aumento de productos de glicosilación, etc) que hacen necesarios cada vez mayores niveles de insulina para conseguir los mismos efectos sobre el control de la glucemia, lo que puede dar lugar a la aparición de resistencia a la insulina.
Cuando se produce una ingesta excesiva a lo largo de los años tiene lugar una serie de consecuencias negativas; así, por ejemplo, las grandes hiperglucemias seguidas de hipoglucemias posprandiales pronunciadas alteran los ritmos de hambre y saciedad, obligando a comer continuamente y desarrollando un apetito dirigido a los productos de mayor densidad calórica y altos en hidratos de carbono refinados. La conjunción de éste y otros factores (sedentarismo, obesidad) dan lugar a alteraciones a nivel celular (reducción de transportadores de glucosa GLUT4, aumento de productos de glicosilación, etc) que hacen necesarios cada vez mayores niveles de insulina para conseguir los mismos efectos sobre el control de la glucemia, lo que puede dar lugar a la aparición de resistencia a la insulina.
Otras hormonas implicadas en el metabolismo de los hidratos de carbono
En términos globales podemos decir que la insulina fomenta el empleo de hidratos de carbono como fuente de energía a la vez que impide el uso de los lípidos. Si la glucemia se eleva se secreta insulina, los hidratos de carbono se utilizan con fines energéticos y su exceso se almacena en forma de glucógeno y grasa; si la glucemia desciende, las células utilizarán las grasas como fuente de energía. Pero además, es una hormona imprescindible para el correcto metabolismo de grasas y proteínas.
Mecanismo de acción de la insulina y alteraciones
Ante una comida con una gran cantidad de hidratos de carbono se libera una gran cantidad de insulina con el objetivo de regular la glucemia sanguínea. La rápida llegada de insulina al torrente sanguíneo permite la entrada de glucosa en las células para ser utilizada como combustible y almacenarse en forma de glucógeno. Llegados a este punto es necesario aclarar (que no desmentir) un mito bastante extendido: "el exceso de carbohidratos de almacena en forma de grasa". Es cierto, como hemos explicado, que una vez se ha superado la capacidad del hígado para almacenar glucógeno, la glucosa puede transformarse en ácidos grasos (lipogénesis) para su reserva en el tejido adiposo; sin embargo, este es un proceso relativamente lento, necesitándose ingestas muy abundantes e incluso más de una comida para que comience este proceso. Facilitan la lipogénesis algunos factores, como la dieta alta en hidratos de carbono de forma crónica (pues los depósitos de glucógeno ya estarán llenos previamente) o el consumo de altas cantidades de fructosa (que solo puede almacenarse como glucógeno hepático pero no muscular).
Ya hemos visto que la insulina se secreta con el objetivo principal de controlar los niveles de glucemia, pero ni son los carbohidratos el único macronutriente que estimula la secreción de insulina, ni la función de ésta es únicamente el manejo de los hidratos de carbono. Comidas que contienen baja cantidad de hidratos de carbono también provocan la secreción de insulina, concepto importante y que tenemos que tener claro para no caer en la falsa creencia de que "la insulina nos hace engordar". Se han estudiado los efectos de un gran número de alimentos sobre la secreción de insulina, descubriéndose que algunas sustancias como la carne de res, el suero de leche (whey), o incluso el aminoácido leucina provocan elevaciones en la secreción de insulina que pueden llegar a alcanzar la misma magnitud que se produciría con el consumo de hidratos de carbono.
Parece entonces poco lógico decir que la insulina es causante de obesidad o que que mantener niveles bajos de insulina nos permitiría adelgazar, y que el consumo crónico y por encima de los límites de tolerancia de cada persona de hidratos de carbono juega un papel más importante en este fenómeno.
En individuos sanos existen dos ritmos de secreción de insulina que se superponen: uno basal, que regula constantemente los niveles de glucemia; y uno en forma de picos como respuesta a las comidas, que facilita la utilización de los nutrientes ingerido; la producción de estos picos de insulina es una situación fisiológica, y es imprescindible para la correcta regulación del metabolismo energético. En personas sanas se repite un ciclo constante de hiperglucemia - hiperinsulinemia - hipoglucemia reactiva a lo largo del día, cuyas oscilaciones serán tanto más pronunciadas conforme mayor sea la cantidad de hidratos de carbono que se consume.
Otras hormonas implicadas en el metabolismo de los hidratos de carbono
El metabolismo energético está ampliamente controlado, y existen otras cuatro hormonas que actúan en equilibrio con la insulina para este propósito. El glucagón induce la glucogenolisis y la gluconeogénesis en situaciones de hipoglucemia. La hormona del crecimiento (GH) y el cortisol aumentan sus niveles en situaciones de hipoglucemia, estimulando el metabolismo lipídico. La adrenalina, por su parte, eleva la glucosa en periodos de estrés, pero también incrementa las concentraciones de ácidos grasos libres.
Glucagón
Glucagón
Entre comidas, la falta de insulina y el aumento de glucagón facilitan la disposición de unidades de glucosa que puedan ser utilizadas como fuente de energía. El glucagón fomenta la glucogenolisis y la gluconeogénesis, empleando aminoácidos cuando ya no queda glucógeno disponible. Aunque clásicamente se ha señalado su efecto estimulador de la lipasa en las células del tejido adiposo proveyendo ácidos grasos para su degradación, éste es un efecto que actualmente ha sido puesto en duda.
Debe existir un equilibrio constante entre insulina y glucagón, de modo que los niveles de glucemia se mantengan aproximadamente constantes, evitando picos excesivos y caídas bruscas.
Debe existir un equilibrio constante entre insulina y glucagón, de modo que los niveles de glucemia se mantengan aproximadamente constantes, evitando picos excesivos y caídas bruscas.
Hormona del crecimiento
Insulina y GH actúan de forma sinérgica favoreciendo el crecimiento celular, aunque no lo hacen de forma sincrónica. La hormona del crecimiento es secretada por la hipófisis anterior en forma de picos y siguiendo un ritmo circadiano con una mayor tasa de producción durante el sueño. Actúa sobre el hígado para producir IGF-1 (factor de crecimiento insulínico), molécula efectora de las acciones de la GH, que permite el desarrollo y crecimiento del tejido muscular, el cartílago, los huesos, la piel, las neuronas y las células sanguíneas entre otros. Esta hormona es estructuralmente similar a la insulina y actúa sobre sus mismos receptores, pero se produce en circunstancias diametralmente opuestas, es decir, cuando los niveles de glucosa en sangre son bajos.
Por tanto, para que ambas hormonas puedan llevar a cabo sus funciones de la forma más eficiente posible resulta imprescindible la diferenciación de dos estados metabólicos: la abundancia y la escasez de glucosa; y se entiende que en un escenario con altas concentraciones de glucosa e insulina se produzca una menor actividad de la GH (como puede ocurrir en personas obesas).
Por tanto, para que ambas hormonas puedan llevar a cabo sus funciones de la forma más eficiente posible resulta imprescindible la diferenciación de dos estados metabólicos: la abundancia y la escasez de glucosa; y se entiende que en un escenario con altas concentraciones de glucosa e insulina se produzca una menor actividad de la GH (como puede ocurrir en personas obesas).
Cortisol
El cortisol es una hormona producida por la corteza suprarrenal y que juega un papel importante en situaciones de estrés. Sus múltiples funciones van encaminadas a la adaptación a situaciones estresantes, como un descanso insuficiente, una actividad física excesiva, o una hipoglucemia. Actúa sobre el metabolismo de hidratos de carbono, grasas y proteínas; y aumenta la disponibilidad de glucosa. Su secreción sigue también un ritmo circadiano, con un pico en los momentos previos al despertar, por lo que cumpliría una función de "despertador fisiológico". El estrés excesivo y los niveles continuamente altos de glucosa en sangre pueden alterar el ritmo de secreción de cortisol, dando lugar a patrones de producción disfuncionales o en cantidades excesivas.
Catecolaminas
Las catecolaminas (adrenalina, noradrenalina) son moléculas producidas por la médula suprarrenal, y regulan los ajustes rápidos a situaciones de estrés, modificando múltiples ritmos y funciones corporales.
En cuanto al metabolismo energético, la acción global de la adrenalina consiste en aumentar la disponibilidad de sustratos energéticos llevando a cabo una serie de acciones como el aumento de la glucogenolisis muscular y la lipogénesis en el tejido adiposo, la disminución de la producción de insulina y el aumento de la producción de glucagón.
Bibliografía
Capítulo 67. Metabolismo de los hidratos de carbono y formación de trifosfato de adenosina. En: Guyton y Hall. Tratado de fisiología médica. 12ª Edición. Elsevier Health Sciences, Aug 30, 2011
Capítulo 14. Glucolisis, gluconeogénesis y vía de las pentosas fosfato. En: Lehninger. Principios de bioquímica. Nelson D. L., Cox M. M. 4ª Edición. Omega, 2006
A.H. Romano, T. Conway. Evolution of carbohydrate metabolic pathways. Research in Microbiology. Volume 147, Issues 6–7, July–September 1996, Pages 448–455
Capítulo 78. Insulina, glucagón y diabetes mellitus. En: Guyton y Hall. Tratado de fisiología médica. 12ª Edición. Elsevier Health Sciences, Aug 30, 2011
Holt SH1, Miller JC, Petocz P. An insulin index of foods: the insulin demand generated by 1000-kJ portions of common foods. Am J Clin Nutr. 1997 Nov;66(5):1264-76.
Yang J, Chi Y, Burkhardt BR, Guan Y, Wolf BA. Leucine metabolism in regulation of insulin secretion from pancreatic beta cells. Nutr Rev. 2010 May;68(5):270-9.
Sener A, Malaisse WJ. The stimulus-secretion coupling of amino acid-induced insulin release: insulinotropic action of branched-chain amino acids at physiological concentrations of glucose and glutamine. Eur J Clin Invest. 1981 Dec;11(6):455-60.
Chaput JP, Tremblay A. The glucostatic theory of appetite control and the risk of obesity and diabetes. Int J Obes (Lond). 2009 Jan;33(1):46-53.
Gravholt CH, Møller N, Jensen MD, Christiansen JS, Schmitz O. Physiological levels of glucagon do not influence lipolysis in abdominal adipose tissue as assessed by microdialysis. J Clin Endocrinol Metab. 2001 May;86(5):2085-9.
Lanzi R, Luzi L, Caumo A, Andreotti AC, Manzoni MF, Malighetti ME, Sereni LP, Pontiroli AE. Elevated insulin levels contribute to the reduced growth hormone (GH) response to GH-releasing hormone in obese subjects. Metabolism. 1999 Sep;48(9):1152-6.
Como siempre, lectura fácil y muy comprensiva.!!!
ResponderEliminaruna cuestión, entonces introducir más de 3 comidas al día, puede producir picos de insulina y a su vez, puede provocar que la perdida de grasa sea mas lenta?
En cetosis o cuando pretendes keto-adaptar se puede meter truvia como sustituto de eritritol¿? he leido que es lo mismo, que opinión tienes al respecto¿?
La secreción de insulina en picos pronunciados no se relacionaría tanto con el número de comidas sino la cantidad de comida y más concretamente de azúcares refinados que contiene la misma. El número de comidas que hacemos puede obedecer a muchos factores, por ejemplo, que necesitemos 4, 5 o incluso más comidas porque nuestras necesidades calóricas son realmente altas (atletas de competición por ejemplo). Lo perjudicial para la acción de la insulina es que, al incluir gran cantidad de azúcares provoquemos continuamente subidas y bajas bruscas de glucemia.
EliminarSobre cómo afecta esto a la pérdida de grasa, siempre hay que relativizar. En el momento inmediatamente posterior a la comida el balance del metabolismo favorece el anabolismo y por tanto no es posible quemar grasa. Pero el balance global a lo largo del día puede favorecer justo lo contrario, por ejemplo porque estamos en dieta hipocalórica, la pérdida de grasa no tiene por qué ralentizarse.
Truvia es básicamente eritritol con un porcentaje marginal de glucósidos de stevia, así que en realidad es lo mismo. Pero si lo piensas bien, la stevia se vende mucho más cara, por lo que creo que económicamente deberías tenerlo en cuenta.
Buen artículo!
ResponderEliminarEn estos momentos estoy probando una dieta low carbo (casi cetogénica) junto a entrenamientos cardio en ayunas en zona fatmax(+0- 65% f. Máx) para mejorar el consumo de grasas. Tengo entendido que de esta manera no hay secreción de insulina y se consumen mayoritariamente grasas.
Que opinas? Eres papartidario?
Gracias y saludos!
Hola Nacho
EliminarInsulina vas a tener siempre, pues como explico en el artículo, siempre hay una secreción basal; lo que evitarías serán los grandes picos que puedan provocar las comidas.
De este modo, vas a quemar grasa durante más tiempo. Así que sí, me parece correcto.